
脳の海馬歯状回の新しい神経細胞が記憶の忘却を促進することを発見 — 幼児期健忘の脳内メカニズムの解明に前進—
システム医科学研究部門の共同研究の成果が Science に掲載されました。
読売新聞、日本経済新聞、CBC、The New York Times、Washington Post、VOX、The Scientist、France inter、Le Sciencze、forskning.noなどをはじめとして世界各国で報道されました。
論文の社会的インパクトを数値化するAltmetric解析では スコア 330 であり、これはトラック可能な 2,128,157 の論文中上位 1% に位置しています(5月19日時点)。
また、ScienceNow、掲載号のPERSPECTIVE、Nature Newsでも取り上げられました。
論文の社会的インパクトを数値化するAltmetric解析では スコア 330 であり、これはトラック可能な 2,128,157 の論文中上位 1% に位置しています(5月19日時点)。
また、ScienceNow、掲載号のPERSPECTIVE、Nature Newsでも取り上げられました。
トロント大学およびトロント小児病院の Paul Frankland 教授らと藤田保健衛生大学総合医科学研究所システム医科学研究部門の宮川剛教授らの日加共同研究チームは、脳の中で学習・記憶に重要な役割を果たす海馬(注1)の歯状回(注2)という領域において、ランニングや抗うつ薬の投与などにより新しい未成熟な神経細胞を増やすと記憶の忘却が促進されることを明らかにしました。本研究の成果は、幼児期健忘(注3)の脳内メカニズムの解明に大きく貢献することが期待されます。この研究成果は、5月9日(日本時間)付けの米科学誌 Science(サイエンス)に、Research Articleとして掲載されました。
研究の背景と経緯
私たちヒトは3歳頃より前に経験した出来事をほとんど思い出すことができません。この現象を「幼児期健忘」と言いますが、そのメカニズムは長い間わかっていませんでした。今回、私たちの研究チームは、幼児期健忘が生じるメカニズムとして、脳の海馬にある歯状回という領域で新たに生まれる未成熟な神経細胞の役割に着目しました。海馬は、学習・記憶をはじめ、情動やストレス反応などの調節に重要であり、近年では精神疾患との関係から注目を集めている領域です(Leuner & Gould, Annual Review of Psychology, 2010)。このような海馬の一部を構成する歯状回では、生涯にわたって新しい神経細胞が生まれること(神経新生(注4))が知られています(Ming & Song, Annual Review of Neuroscience, 2005; Zhao et al., Cell, 2008)。新生した神経細胞は、はじめは未成熟な状態ですが、やがて成熟し、既存の神経回路に組み込まれていきます(Toni et al., Nature Neuroscience, 2008; Zhao et al., Journal of Neuroscience, 2006; Gu et al., Nature Neuroscience, 2012)。これまでの研究から、海馬歯状回の神経新生は新しい記憶の形成に重要な役割を果たしていることが報告されてきました(Sahay et al., Nature, 2011; Stone et al., Journal of Neuroscience, 2011)。その一方で、神経新生が起こると、既存の神経回路に再編が生じるため、それまで海馬の神経回路に蓄えられていた記憶は回路の再編に伴って忘却されてしまう可能性があるという指摘もあります(Meltzer et al., Trends in Neuroscience, 2005; Weisz & Argibay, Cognition, 2012)。歯状回での神経新生は個体の成熟過程とともに減少することが知られていますが(Kuhn et al., Journal of Neuroscience, 1996; Seki & Arai, Neuroreport, 1995)、もし神経新生により記憶の忘却が促進されるのであれば、神経新生が活発な子供の時に経験した出来事の記憶は忘却されやすく、神経新生が少ない大人の時に経験した出来事の記憶は忘却されにくいと考えられます。そこで、本研究では、子供と大人のマウスあるいはデグーなどのげっ歯類を用いて、行動神経科学的・行動薬理学的手法および遺伝子改変技術などを駆使し、海馬歯状回での神経新生と記憶の忘却との関係について調べました。
研究の内容
本研究では、はじめに子供のマウスと大人のマウスの歯状回における神経新生の違いを比較しました。新生した神経細胞は蛍光を発するように標識して可視化しました。子供のマウスでは、大人のマウスと比べて、歯状回で新しい神経細胞が盛んに産生されていることと、歯状回—CA3領域の神経回路において新しい神経細胞由来の神経終末が多く存在していることを確認しました。これらのことは、子供のマウスの海馬では、大人と比較して、神経回路の改編が盛んにおこっていることを示しています(図1—(1))。
続いて、子供と大人のマウスの記憶能力を評価しました。ある環境にマウスを置いて、足に軽い電気ショックを与えると、マウスはその環境と電気ショックとの関係を学習します。再びマウスをその環境におくと、「この場所で電気ショックを受けた」という恐い記憶を思い出して身動きしない反応(すくみ反応)をするようになります。このすくみ反応を示した時間の長さを測定することによって、子供と大人のマウスの記憶能力について調べることができます(恐怖条件づけテスト(注5))。大人のマウスは学習してから4週間後にテストを行っても学習後1日とほとんど変わらない時間のすくみ反応を示しますが、子供のマウスでは学習してから1週間後にテストを行った時点で、すでにすくみ反応の時間が短くなっており、子供のマウスのほうが記憶の忘却が起こりやすいことがわかりました(図1—(2))。これらの結果は、ヒトの幼児性健忘と同様の現象がマウスでも起きていることを示唆しています。
続いて、子供と大人のマウスの記憶能力を評価しました。ある環境にマウスを置いて、足に軽い電気ショックを与えると、マウスはその環境と電気ショックとの関係を学習します。再びマウスをその環境におくと、「この場所で電気ショックを受けた」という恐い記憶を思い出して身動きしない反応(すくみ反応)をするようになります。このすくみ反応を示した時間の長さを測定することによって、子供と大人のマウスの記憶能力について調べることができます(恐怖条件づけテスト(注5))。大人のマウスは学習してから4週間後にテストを行っても学習後1日とほとんど変わらない時間のすくみ反応を示しますが、子供のマウスでは学習してから1週間後にテストを行った時点で、すでにすくみ反応の時間が短くなっており、子供のマウスのほうが記憶の忘却が起こりやすいことがわかりました(図1—(2))。これらの結果は、ヒトの幼児性健忘と同様の現象がマウスでも起きていることを示唆しています。
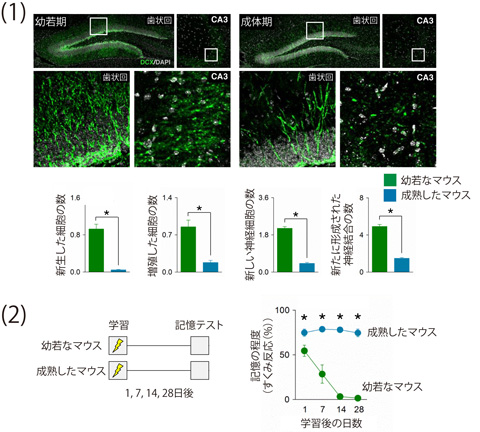
図1 幼若期と成体期における海馬歯状回の神経細胞の新生と記憶の忘却の関係
(1)子供のマウスの歯状回では、大人のマウスに比べて、新しい神経細胞(緑色)がたくさん生まれ、CA3領域との間に新しい神経結合が形成されていた。
(2)ある環境で軽い電気ショックを受けた大人のマウスは、再びその環境下に置かれるとすくみ反応を示すが、この反応は4週間後もほぼ同じ時間維持されていた。つまり、過去の嫌な出来事の記憶を4週間後も保持していたことを意味している。一方、子供のマウスでは、電気ショックを受けてから1週間以上経過すると、すくみ反応の時間が短くなっていた。この結果は、子供のマウスでは記憶の忘却が起きていることを示唆している。
(1)子供のマウスの歯状回では、大人のマウスに比べて、新しい神経細胞(緑色)がたくさん生まれ、CA3領域との間に新しい神経結合が形成されていた。
(2)ある環境で軽い電気ショックを受けた大人のマウスは、再びその環境下に置かれるとすくみ反応を示すが、この反応は4週間後もほぼ同じ時間維持されていた。つまり、過去の嫌な出来事の記憶を4週間後も保持していたことを意味している。一方、子供のマウスでは、電気ショックを受けてから1週間以上経過すると、すくみ反応の時間が短くなっていた。この結果は、子供のマウスでは記憶の忘却が起きていることを示唆している。
次に、歯状回の神経新生と記憶の忘却の因果関係を明確にするために、大人のマウスを用いて、神経細胞の新生を高めることが可能な各種実験操作を行い、記憶の忘却が促進されるかどうかを調べました。大人のマウスにランニング(注6)をさせたり、アルツハイマー型認知症の治療薬として知られるメマンチン(NMDA受容体拮抗薬)(注7)や抗うつ薬であるフルオキセチン(選択的セロトニン再取り込み阻害薬)(注8)を投与すると、神経新生を増やすことができます。学習してから3週から6週の間、それらの方法によって歯状回の神経新生を増やした大人のマウスでは、何もしなかった場合に比べて、記憶の忘却が促進されることを見出しました(図2、図3)。一方、歯状回の神経新生が活発な子供のマウスに神経新生を抑制することのできる遺伝子操作や薬物投与を行うと、記憶の忘却が抑制されることがわかりました。また、同じげっ歯類でも、マウスやラットと異なり、モルモットやデグーは、妊娠期間が長く、出生した時点で既に毛が生え、目が開いているなど、ある程度発達した状態で生まれてきます(早成性(注9))。このような早成性げっ歯類の子供は、大人と比べても、歯状回での神経新生の程度に大きな違いがなく記憶の忘却の程度もほとんど差が見られませんでしたが、ランニングや薬物投与によって早成性げっ歯類の子供の神経新生を増やすと記憶の忘却が促進されることがわかりました。
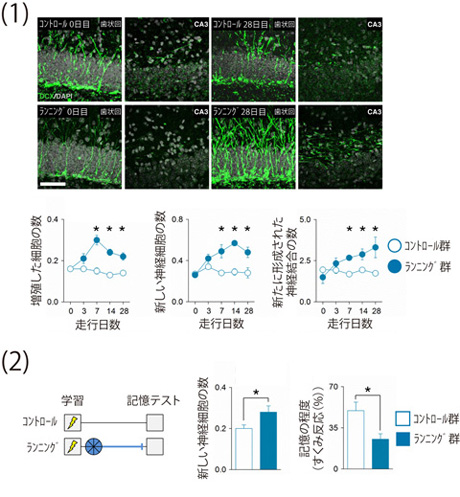
図2 ランニングによる海馬歯状回の神経細胞の新生と記憶の忘却の促進
(1)飼育ケージに回転カゴを設置すると、マウスはそのカゴの中に入り、自発的にランニングをするようになる。大人のマウスにランニングをする機会を与えてから1週間以上経過すると、そのマウスの歯状回では新しい神経細胞がたくさん産生されていた。
(2)大人のマウスは、学習してから記憶テストを受けるまでの6週間の間、飼育ケージ内にある回転カゴの中で自由にランニングをする機会が与えられると、記憶の低下を示した。
(1)飼育ケージに回転カゴを設置すると、マウスはそのカゴの中に入り、自発的にランニングをするようになる。大人のマウスにランニングをする機会を与えてから1週間以上経過すると、そのマウスの歯状回では新しい神経細胞がたくさん産生されていた。
(2)大人のマウスは、学習してから記憶テストを受けるまでの6週間の間、飼育ケージ内にある回転カゴの中で自由にランニングをする機会が与えられると、記憶の低下を示した。
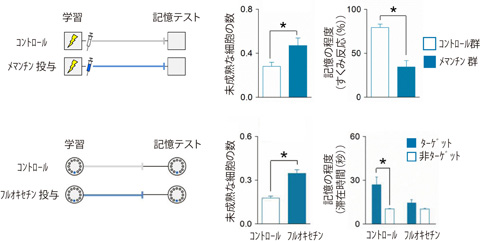
図3 薬物投与による海馬歯状回の神経細胞の新生と記憶の忘却の促進
メマンチン(NMDA受容体拮抗薬)やフルオキセチン(選択的セロトニン再取り込み阻害薬)を投与することにより、歯状回において新しい未成熟な神経細胞を増加させると、記憶の忘却が促進された(上図;恐怖条件づけテスト、下図;空間参照記憶テスト)。
メマンチン(NMDA受容体拮抗薬)やフルオキセチン(選択的セロトニン再取り込み阻害薬)を投与することにより、歯状回において新しい未成熟な神経細胞を増加させると、記憶の忘却が促進された(上図;恐怖条件づけテスト、下図;空間参照記憶テスト)。
これまでに神経新生と記憶の関係を調べるために行われた研究の多くは、神経新生を誘導してからマウスに学習させ、記憶能力をテストするという手続きを用いていました(Deng et al., Nature Reviews Neuroscience, 2010)。その結果から導きだれた結論は、神経新生は新しい記憶の形成に重要であるというものでした。今回、私たちが用いた手続きは、学習後に神経新生を誘導し、記憶能力をテストするというものです。これにより、神経新生が既に獲得した記憶の忘却を促進するという現象を発見し、神経新生の新たな役割を明らかにすることができました。以上の結果から、幼若期に海馬歯状回において盛んに生じている神経新生によって記憶の忘却が促進され、神経細胞の新生が少ない成体期には記憶の忘却が起こりにくいということが明らかになりました。これらの知見は、海馬歯状回で盛んに産生される新しい未成熟な神経細胞が幼児期健忘を引き起こすメカニズムとして重要であることを示しています。
今後の展開
本研究により、海馬歯状回で新しく生まれた未成熟な神経細胞が幼児期健忘に関与していることが明らかとなりました。しかしながら、新生した神経細胞がどのようにして記憶の忘却を促進しているのかについてはまだほとんど明らかになっていません。そのため、分子レベルおよび神経回路レベルにおいて、記憶の忘却を引き起こすメカニズムの解明に向けた研究を進めていく必要があります。また、本研究の成果は、げっ歯類を対象に行った実験から得られたものであるため、実際にヒトの幼児期健忘が同様のメカニズムで生じているのかについて検討する必要があります。また、大人のマウスで歯状回の神経新生を促進させると記憶の忘却が促進されたことから、今後、ヒトを含めた哺乳類における歯状回の神経新生による記憶の忘却についての分子および回路レベルのメカニズムをさらに解明することで、強いショック体験や精神的ストレスが原因となって生じる心的外傷後ストレス障害(PTSD)などの精神疾患の病態理解および予防・治療法の開発や創薬につながることが期待されます。
参考文献
Deng, W., Aimone, J.B., Gage, F.H. (2010). New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nature Reviews Neuroscience, 11, 339–350.
Frankland, P.W., Köhler, S., Josselyn, S.A. (2013). Hippocampal neurogenesis and forgetting. Trends in Neuroscience, 36, 497–503.
Gu, Y., Arruda-Carvalho, M., Wang, J., Janoschka, S.R., Josselyn, S.A., Frankland, P.W., Ge, S. (2012). Optical controlling reveals time-dependent roles for adult-born dentate granule cells. Nature Neuroscience, 15, 1700–1706.
Kobayashi, K., Ikeda, Y., Sakai, A., Yamasaki, N., Haneda, E., Miyakawa, T., Suzuki, H. (2010). Reversal of hippocampal neuronal maturation by serotonergic antidepressants. Proceedings of the National Academy of Sciences of the United States of America, 107, 8434–8439.
Kuhn, H.G., Dickinson-Anson, H., Gage, F.H. (1996). Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. Journal of Neuroscience, 16, 2027–2033.
Leuner, B., Gould, E. (2010). Structural plasticity and hippocampal function. Annual Review of Psychology, 61, 111-140.
Malberg, J.E., Eisch, A.J., Nestler, E.J., Duman, R.S. (2000). Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. Journal of Neuroscience, 20, 9104–9110.
Meltzer, L.A., Yabaluri, R., Deisseroth, K. (2005). A role for circuit homeostasis in adult neurogenesis. Trends in Neuroscience, 28, 653–660.
Ming, G.L., Song, H. (2005). Adult neurogenesis in the mammalian central nervous system. Annual Review of Neuroscience, 28, 223–250.
Ohira, K., Takeuchi, R., Shoji, H., Miyakawa, T. (2013). Fluoxetine-induced cortical adult neurogenesis. Neuropsychopharmacology, 38, 909–920.
Sahay, A., Scobie, K.N., Hill, A.S., O'Carroll, C.M., Kheirbek, M.A., Burghardt, N.S., Fenton, A.A., Dranovsky, A., Hen, R. (2011). Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature, 472, 466–470.
Seki, T., Arai, Y. (1995). Age-related production of new granule cells in the adult dentate gyrus. Neuroreport, 6, 2479–2482.
Stone, S.S., Teixeira, C.M., DeVito, L.M., Zaslavsky, K., Josselyn, S.A., Lozano, A.M., Frankland, P.W. (2011). Stimulation of entorhinal cortex promotes adult neurogenesis and facilitates spatial memory. Journal of Neuroscience, 31, 13469–13484.
Toni, N., Laplagne, D.A., Zhao, C., Lombardi, G., Ribak, C.E., Gage, F.H., Schinder, A.F. (2008). Neurons born in the adult dentate gyrus form functional synapses with target cells. Nature Neuroscience, 11, 901–907.
Weisz, V.I., Argibay, P.F. (2012). Neurogenesis interferes with the retrieval of remote memories: forgetting in neurocomputational terms. Cognition, 125, 13–25.
Zhao, C., Deng, W., Gage, F.H. (2008). Mechanisms and functional implications of adult neurogenesis. Cell, 132, 645–660.
Zhao, C., Teng, E.M., Summers, R.G., Ming, G.L., Gage, F.H. (2006). Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. Journal of Neuroscience, 26, 3–11.
Frankland, P.W., Köhler, S., Josselyn, S.A. (2013). Hippocampal neurogenesis and forgetting. Trends in Neuroscience, 36, 497–503.
Gu, Y., Arruda-Carvalho, M., Wang, J., Janoschka, S.R., Josselyn, S.A., Frankland, P.W., Ge, S. (2012). Optical controlling reveals time-dependent roles for adult-born dentate granule cells. Nature Neuroscience, 15, 1700–1706.
Kobayashi, K., Ikeda, Y., Sakai, A., Yamasaki, N., Haneda, E., Miyakawa, T., Suzuki, H. (2010). Reversal of hippocampal neuronal maturation by serotonergic antidepressants. Proceedings of the National Academy of Sciences of the United States of America, 107, 8434–8439.
Kuhn, H.G., Dickinson-Anson, H., Gage, F.H. (1996). Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. Journal of Neuroscience, 16, 2027–2033.
Leuner, B., Gould, E. (2010). Structural plasticity and hippocampal function. Annual Review of Psychology, 61, 111-140.
Malberg, J.E., Eisch, A.J., Nestler, E.J., Duman, R.S. (2000). Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. Journal of Neuroscience, 20, 9104–9110.
Meltzer, L.A., Yabaluri, R., Deisseroth, K. (2005). A role for circuit homeostasis in adult neurogenesis. Trends in Neuroscience, 28, 653–660.
Ming, G.L., Song, H. (2005). Adult neurogenesis in the mammalian central nervous system. Annual Review of Neuroscience, 28, 223–250.
Ohira, K., Takeuchi, R., Shoji, H., Miyakawa, T. (2013). Fluoxetine-induced cortical adult neurogenesis. Neuropsychopharmacology, 38, 909–920.
Sahay, A., Scobie, K.N., Hill, A.S., O'Carroll, C.M., Kheirbek, M.A., Burghardt, N.S., Fenton, A.A., Dranovsky, A., Hen, R. (2011). Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature, 472, 466–470.
Seki, T., Arai, Y. (1995). Age-related production of new granule cells in the adult dentate gyrus. Neuroreport, 6, 2479–2482.
Stone, S.S., Teixeira, C.M., DeVito, L.M., Zaslavsky, K., Josselyn, S.A., Lozano, A.M., Frankland, P.W. (2011). Stimulation of entorhinal cortex promotes adult neurogenesis and facilitates spatial memory. Journal of Neuroscience, 31, 13469–13484.
Toni, N., Laplagne, D.A., Zhao, C., Lombardi, G., Ribak, C.E., Gage, F.H., Schinder, A.F. (2008). Neurons born in the adult dentate gyrus form functional synapses with target cells. Nature Neuroscience, 11, 901–907.
Weisz, V.I., Argibay, P.F. (2012). Neurogenesis interferes with the retrieval of remote memories: forgetting in neurocomputational terms. Cognition, 125, 13–25.
Zhao, C., Deng, W., Gage, F.H. (2008). Mechanisms and functional implications of adult neurogenesis. Cell, 132, 645–660.
Zhao, C., Teng, E.M., Summers, R.G., Ming, G.L., Gage, F.H. (2006). Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. Journal of Neuroscience, 26, 3–11.
用語解説
(注1)海馬
海馬は、大脳辺縁系の一部で、学習・記憶に重要な役割を果たしています。CA1、CA2、CA3、歯状回の各部位から構成されています。
(注2)歯状回
歯状回は、海馬の一領域で、学習・記憶などの認知機能との関連のほか、近年は精神疾患との関連で注目されています。
(注3)幼児期健忘
ヒトは乳幼児の頃の記憶を覚えていないものですが、心理学ではこれを幼児期健忘といいます。健忘を生じる理由として、乳幼児期は未熟で記憶をうまく固定できないという考えや、記憶を貯蔵する際に必要であった神経回路が発達する過程で新たに形成された神経回路に飲み込まれてしまい,当時の記憶を思い出せないという考えが提唱されていました。
(注4)神経新生
大人の脳では海馬歯状回と脳室周囲に神経幹細胞が存在し、神経幹細胞が多数分裂して数を増やし、神経細胞やグリア細胞に分化します。この過程を「神経新生」と呼んでいます。神経新生は、加齢やストレスにより減少し、ランニングや薬物摂取によって増加することが知られています。
(注5)恐怖条件づけテスト
マウスやラットの恐怖に関連する記憶を測定するテスト。マウスやラットは、ある環境下で軽い電気ショックを受けると、その環境と電気ショックとの関係を学習し、その後、同じ環境にさらされると恐怖によるすくみ反応を示します。すくみ反応を示した時間が記憶能力の指標として用いられています。
(注6)ランニング
飼育ケージに回転かごを設置すると、ネズミはカゴの中に入って自発的に走るようになります。実験動物のマウスは、一日あたりおよそ5kmの距離を走行します。
(注7)メマンチン
欧米で承認されているアルツハイマー型認知症の治療薬です。NMDA型グルタミン酸受容体の働きを抑制することによって脳の神経細胞の過剰な興奮による細胞死を防ぎます。このメマンチンを投与すると、神経新生が増加することが報告されています。
(注8)フルオキセチン
フルオキセチンは、選択的セロトニン再取り込み阻害薬という抗うつ薬の一つです。セロトニンは、生理活性アミンの一種で、生体リズムや睡眠、気分障害や統合失調症などの病態に関与しています。フルオキセチンは、神経終末から放出されたセロトニンが神経末端に取り込まれることを防ぎ、細胞外のセロトニン濃度を上昇させます。フルオキセチンの長期投与により、歯状回での神経新生が増加すること、歯状回をはじめ扁桃体や前頭皮質の特定の神経細胞が成熟した状態から未成熟な状態に戻ることが知られています。
(注9)早成性
動物が生後まもなくから体温維持能力や移動能力を有するなど、自立して生活できる特徴のことを指します。これとは対照的に、ヒトやサル、マウスなどは、生後しばらくの間は未熟で養育者からの保護を必要とする特徴を示します(晩生性)。
海馬は、大脳辺縁系の一部で、学習・記憶に重要な役割を果たしています。CA1、CA2、CA3、歯状回の各部位から構成されています。
(注2)歯状回
歯状回は、海馬の一領域で、学習・記憶などの認知機能との関連のほか、近年は精神疾患との関連で注目されています。
(注3)幼児期健忘
ヒトは乳幼児の頃の記憶を覚えていないものですが、心理学ではこれを幼児期健忘といいます。健忘を生じる理由として、乳幼児期は未熟で記憶をうまく固定できないという考えや、記憶を貯蔵する際に必要であった神経回路が発達する過程で新たに形成された神経回路に飲み込まれてしまい,当時の記憶を思い出せないという考えが提唱されていました。
(注4)神経新生
大人の脳では海馬歯状回と脳室周囲に神経幹細胞が存在し、神経幹細胞が多数分裂して数を増やし、神経細胞やグリア細胞に分化します。この過程を「神経新生」と呼んでいます。神経新生は、加齢やストレスにより減少し、ランニングや薬物摂取によって増加することが知られています。
(注5)恐怖条件づけテスト
マウスやラットの恐怖に関連する記憶を測定するテスト。マウスやラットは、ある環境下で軽い電気ショックを受けると、その環境と電気ショックとの関係を学習し、その後、同じ環境にさらされると恐怖によるすくみ反応を示します。すくみ反応を示した時間が記憶能力の指標として用いられています。
(注6)ランニング
飼育ケージに回転かごを設置すると、ネズミはカゴの中に入って自発的に走るようになります。実験動物のマウスは、一日あたりおよそ5kmの距離を走行します。
(注7)メマンチン
欧米で承認されているアルツハイマー型認知症の治療薬です。NMDA型グルタミン酸受容体の働きを抑制することによって脳の神経細胞の過剰な興奮による細胞死を防ぎます。このメマンチンを投与すると、神経新生が増加することが報告されています。
(注8)フルオキセチン
フルオキセチンは、選択的セロトニン再取り込み阻害薬という抗うつ薬の一つです。セロトニンは、生理活性アミンの一種で、生体リズムや睡眠、気分障害や統合失調症などの病態に関与しています。フルオキセチンは、神経終末から放出されたセロトニンが神経末端に取り込まれることを防ぎ、細胞外のセロトニン濃度を上昇させます。フルオキセチンの長期投与により、歯状回での神経新生が増加すること、歯状回をはじめ扁桃体や前頭皮質の特定の神経細胞が成熟した状態から未成熟な状態に戻ることが知られています。
(注9)早成性
動物が生後まもなくから体温維持能力や移動能力を有するなど、自立して生活できる特徴のことを指します。これとは対照的に、ヒトやサル、マウスなどは、生後しばらくの間は未熟で養育者からの保護を必要とする特徴を示します(晩生性)。
掲載論文
この研究成果は、米国科学誌 Science(サイエンス)(日本時間 5月9日付け)に掲載されました。
Katherine G. Akers, Alonso Martinez-Canabal, Leonardo Restivo, Adelaide P. Yiu, Antonietta De Cristofaro, Hwa-Lin (Liz) Hsiang, Anne L. Wheeler, Axel Guskjolen, Yosuke Niibori, Hirotaka Shoji, Koji Ohira, Blake A. Richards, Tsuyoshi Miyakawa, Sheena A. Josselyn & Paul W. Frankland.
Hippocampal neurogenesis regulates forgetting during adulthood and infancy. Science 344: 598-602, 2014.
Katherine G. Akers, Alonso Martinez-Canabal, Leonardo Restivo, Adelaide P. Yiu, Antonietta De Cristofaro, Hwa-Lin (Liz) Hsiang, Anne L. Wheeler, Axel Guskjolen, Yosuke Niibori, Hirotaka Shoji, Koji Ohira, Blake A. Richards, Tsuyoshi Miyakawa, Sheena A. Josselyn & Paul W. Frankland.
Hippocampal neurogenesis regulates forgetting during adulthood and infancy. Science 344: 598-602, 2014.
お問い合わせ先
Paul Frankland
Program in Neurosciences and Mental Health, The Hospital for Sick Children, Toronto, M5G 1X8, Canada.
Tel: +1-416-813-7654 Fax: +1-416-813-7717
Email: paul.frankland[at]sickkids.ca
日本語での問い合わせ先
宮川 剛(みやかわ つよし)
藤田保健衛生大学総合医科学研究所システム医科学 教授
〒470-1192 愛知県豊明市沓掛町田楽ヶ窪1−98
Tel: 0562-93-9375 Fax: 0562-92-5382
Email: miyakawa[at]fujita-hu.ac.jp
Program in Neurosciences and Mental Health, The Hospital for Sick Children, Toronto, M5G 1X8, Canada.
Tel: +1-416-813-7654 Fax: +1-416-813-7717
Email: paul.frankland[at]sickkids.ca
日本語での問い合わせ先
宮川 剛(みやかわ つよし)
藤田保健衛生大学総合医科学研究所システム医科学 教授
〒470-1192 愛知県豊明市沓掛町田楽ヶ窪1−98
Tel: 0562-93-9375 Fax: 0562-92-5382
Email: miyakawa[at]fujita-hu.ac.jp